

Abstract
Background
Following activation by recognition of foreign antigens, human T-cells alter their metabolic pathways to meet the increasing energetic demands for efficient immune response. Like cancer cells, alloreactive T-cells show a preference for aerobic glycolysis rather than oxidative phosphorylation, which is referred to as "Warburg effect". Until recently, it has been thought that extracellular fatty acid (FA) uptake and β-oxidation are severely reduced in alloreactive T-cells; however, some studies have indicated that lipid metabolism is rather increased in alloreactive mouse T-cells, and that metabolic pathway of FA can be a promising target for GVHD. To determine the role of lipid metabolism in human alloreactive T-cells after hematopoietic stem cell transplantation, we investigated the metabolic changes in human T-cells in vivo using human-into-mouse xenogeneic GVHD models.
Methods
NOG mice received 250cGy of total body irradiation (TBI) and were subsequently injected intravenously with human pan T-cells. All mice developed severe GVHD and died within 2 weeks, while mice that received TBI only survived without any symptoms of GVHD. Cells were harvested from GVHD target organs of mice at day 9 after transplantation. For the measurement of glucose and fatty acid (FA) uptake by flow cytometry, cells were stained with fluorescent-labeled deoxyglucose analogue (2-NBDG) and long-chain fatty acid analogue (BODIPY 500/510 C12), respectively. PCR array and extracellular flux analysis were performed according to manufacturer's instructions.
Results
Glucose uptake, determined by flow cytometry, was significantly increased in human T-cells obtained from GVHD mice. Extracellular FA uptake was also increased in human T-cells in GVHD mice, and was associated with cell proliferation rate. Effector memory T-cells followed by central memory T-cells showed a higher FA uptake than did naive T-cells. These findings were similarly observed in both human CD4+ and CD8+ T-cells. Robust T-cell proliferation was observed even in MHC class I/II deficient (MHC−/−) NOG mice after transplantation, although to a lesser extent than MHC+/+ NOG mice, in a process known as homeostatic proliferation. Extracellular uptake of FA as well as glucose in T-cells was significantly decreased in MHC−/− NOG mice. Of note, even when compared among only fully proliferated T-cells between MHC+/+ and MHC−/− NOG mice, FA uptake was still significantly decreased in MHC−/− NOG mice, suggesting that the recognition of host MHC molecules by allogeneic T-cells accelerate this process.
To compare the ability of human naive and memory T-cells to incorporate extracellular FA, we isolated human naive (CD45RA high) and memory (CD45RA low) T-cells and separately injected into NOG mice. Although it has been shown that memory T-cells exhibit different effector functions, the FA uptake in memory T-cells was comparable to that in naive T-cells. This suggests that memory T-cells can also alter their lipid metabolism following encounter with alloantigens.
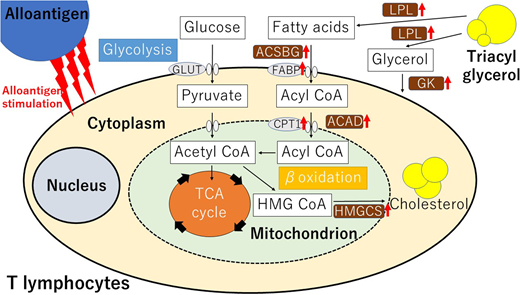
Finally, we assessed the expression of genes associated with lipid metabolism in human T-cells obtained from GVHD mice. Quantitative real-time PCR analysis detected up-regulation of mRNAs encoding the enzymes involved in FA transport including carnitine palmitoyltransferase (CPT1B), fatty acid binding protein (FABP1-4, FABP6, and FABP7), and β-oxidation pathway including acyl-CoA synthase (ACSBG2) and acyl-CoA dehydrogenase (ACAD9-11, ACADS, and ACADL) when compared with T-cells in MHC−/− NOG mice. Similarly, the expression of genes encoding the enzymes in triacylglycerol metabolism such as glycerol kinase (GK, GK2) and lipoprotein lipase (LPL) was up-regulated in GVHD mice. Furthermore, the expression of genes associated with mevalonate pathways such as HMG-CoA synthase (HMGCS1, HMGCS2), was also upregulated. These observations suggest that T-cells activated by alloantigens in vivo promote lipid hydrolysis, mitochondrial FA transport, and β-oxidation, resulting in greater utilization of free FA.
Conclusion
Human alloreactive T-cells increased extracellular uptake of FA as well as glucose, and intracellular lipid metabolism in response to alloantigens (summarized in the graphical abstract). Therapeutic effects of specific inhibition of lipid metabolic pathways by pharmacological inhibitors including etomoxir are now being investigated in this model.
Fujiwara:Shire: Consultancy; Pfizer: Consultancy; Chugai: Consultancy; Kirin: Consultancy; Kyowa-Hakko: Consultancy; Astellas: Consultancy. Ohmine:Kyowa Hakko Kirin: Speakers Bureau; Takara Bio: Membership on an entity's Board of Directors or advisory committees, Research Funding, Speakers Bureau; Takeda Pharmaceutical: Speakers Bureau; Celgene Corporation: Speakers Bureau; Chugai Pharmaceutical: Speakers Bureau; Alexion Pharmaceuticals: Speakers Bureau; Ono Pharmaceutical: Consultancy. Muroi:Japanese Red Cross Society: Speakers Bureau; Dickinson and Company: Speakers Bureau; Becton: Speakers Bureau; JCR: Speakers Bureau. Kanda:Astellas: Consultancy, Honoraria, Research Funding; Eisai: Consultancy, Honoraria, Research Funding; Taiho: Research Funding; Nippon-Shinyaku: Research Funding; Chugai: Consultancy, Honoraria, Research Funding; Dainippon-Sumitomo: Consultancy, Honoraria, Research Funding; Pfizer: Research Funding; Otsuka: Research Funding; Shionogi: Consultancy, Honoraria, Research Funding; Kyowa-Hakko Kirin: Consultancy, Honoraria, Research Funding; MSD: Research Funding; Takeda: Consultancy, Honoraria, Research Funding; Asahi-Kasei: Research Funding; Ono: Consultancy, Honoraria, Research Funding; Sanofi: Research Funding; Novartis: Research Funding; Taisho-Toyama: Research Funding; CSL Behring: Research Funding; Tanabe-Mitsubishi: Research Funding; Bristol-Myers Squibb: Consultancy, Honoraria; Celgene: Consultancy, Honoraria; Mochida: Consultancy, Honoraria; Alexion: Consultancy, Honoraria; Takara-bio: Consultancy, Honoraria.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal